Not surprisingly, this basic idea has been challenged since its inception, and my example for this week comes from another article by Dr. Andrew Snelling of AiG, entitled Order in the Fossil Record. There, Dr. Snelling not only challenges the conventional interpretation that the order of organisms in the fossil record represents the evolutionary history of life, but presents a model employing the Flood as the mechanism for fossil sorting and preservation in the sedimentary rocks that are now a mausoleum of Earth’s past life forms. The best way to do this, he argues, “is to examine a geographic region where the rock layers and fossils are well exposed and well studied. A spectacular example is the Colorado Plateau of the southwestern USA, and more specifically, the Grand Canyon—Grand Staircase rock layers sequence.” If you’ve visited any of the national parks found in this region, then you can certainly appreciate his reasoning. Nearly 3 miles of sedimentary rock layers are found stacked upon one another, and exposed in hundreds of miles of canyons and cliffs throughout northern Arizona and southern Utah. So what do the fossils of these rocks reveal? Before answering that question, let’s consider the Flood geology model.
Flood geology and the Grand Canyon/Grand Staircase sedimentary sequences
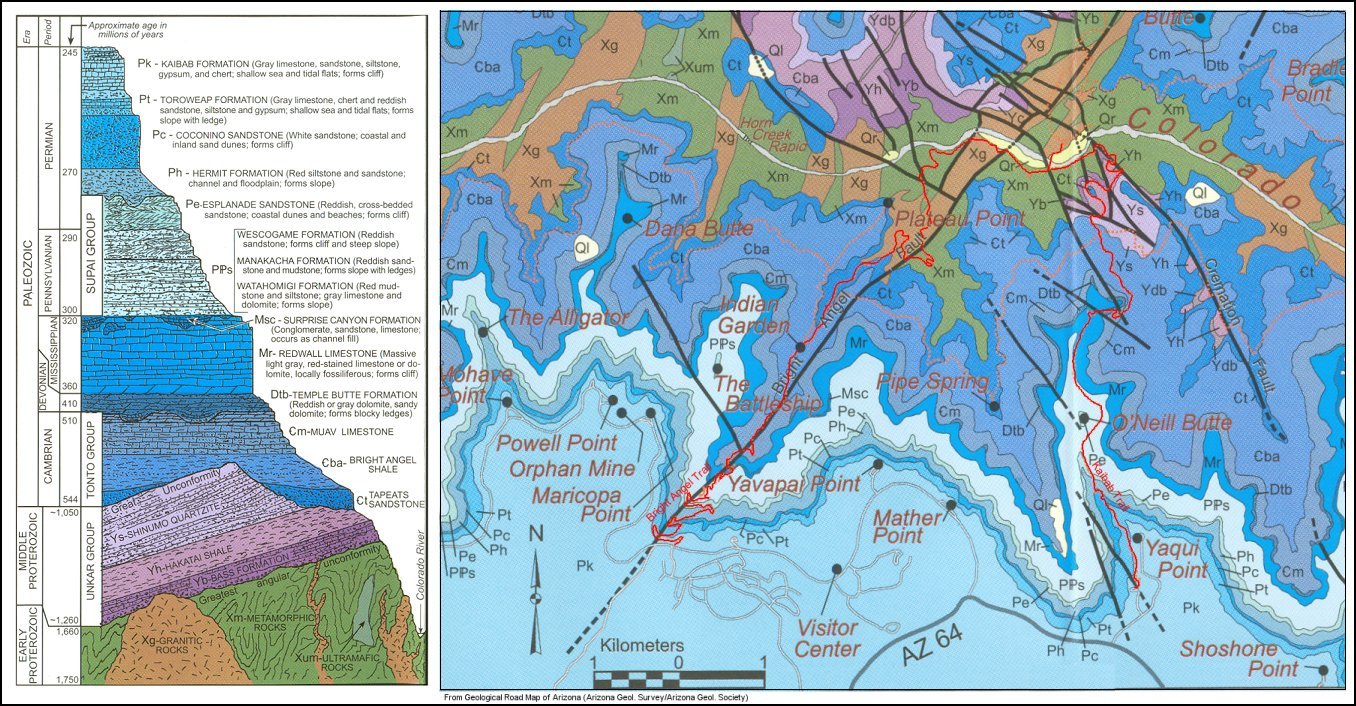
In case you are not familiar with the basic geology of the Grand Canyon, a graphic overview of sedimentary rock layers can be found here, while Dr. Snelling provides a summary of fossils found in the whole sequence here (downloads a 1-page pdf file).
{kind=link}
At the base of the Grand Canyon lies a world famous example of an angular unconformity (see photo here), called the Great Unconformity. This means that sedimentary rocks were deposited in the Precambrian (or before the Flood), hardened into rock, and then tilted upward by tectonic forces before erosion flattened and scoured the surface, onto which the next set of sediments could be deposited. Flood geologists generally interpret the Great Unconformity as an erosive boundary that marks the onset of the Flood (Austin, 1994). It is a regional discontinuity that is also present in southern Nevada and northern Utah, and separates Phanerozoic sedimentary rocks (Cambrian through recent) from underlying Precambrian sedimentary rocks (Unkar and Chuar groups) and crystalline basement rocks (i.e. igneous granites and metamorphic schist bodies). Although numerous fossils are present in the Precambrian Chuar group, they are typically algal in origin (e.g. stromatolites, concretions of algae called acritarchs, etc.), and are thus consistent with the presence of rather calm oceans during the pre-Flood period.
{kind=link}
As the Flood ensued, raging waters rushed over the continent, carrying with them millions of tons of sediment (sand, silt, carbonate mud), which was deposited in the layers seen today. Not surprisingly, a vast majority of organisms living at that time were swept away by the sediment-loaded waters and entombed within the sequence. Tracks of animals desperate to escape (such as trilobites) can be found in the Tapeats Sandstone at the base of the sequence, while their bodies are abundant in the overlying layers. The order of marine fossils thus represents local environments of organisms in approximately the same order they were swept up by the flood. Presumably, rising and falling waters left alternating ‘continental’ deposits, dominated by sandstone and shale that carried land-dwelling organisms, and ‘marine’ deposits, dominated by carbonate mud and shale that carried ocean-dwelling organisms. At the same time, larger, more buoyant animals (such as elephants, horses, etc.) not only had the instincts to escape danger, but were preferentially kept from burial until the end. Thus the order of fossils seen throughout the Grand Canyon Supergroup (and overlying layers in the Grand Staircase) reflects a combination of several mechanisms: 1) ecological zonation, which is determined by the original habitat of the organism (e.g. deep marine, shallow marine, coastal, upland); 2) survivability, which was determined by the organism’s ability to escape catastrophic burial; and 3) hydrodynamic sorting, in which moving waters could sort organisms into different groups based on shape/size.
Precambrian geology of the Grand Canyon
As I began researching this article more in depth, I came to appreciate why most geological research in the Grand Canyon has focused on the Precambrian units (namely, the Unkar Group, Nankoweap Formation, Chuar Group, and Sixtymile Formation). Together, these units comprise some 3,900 meters (that’s 2 1/2 miles) of sedimentary rocks. That’s not including up to 300 meters of lava flows (with interbedded sandstone), and the fact that the rocks were faulted both during and after deposition, which was followed by broad-scale folding. A geologic history longer than the main rocks of the Grand Canyon is buried directly below it, and has conveniently been swept into the “Pre-Flood” category without much consideration. Most of the rocks are marine limestone, dolostone, and shale, indicative of calm tropical seas. Keep in mind that elsewhere, Dr. Snelling proposed that dolomite should not form in vast quantities in the ocean, hence the need for volcanic fluids associated with the Flood as a dolomitizing mechanism. Furthermore, abundant fossils can be found in the sediments of the Chuar Group, yet none of them are multicellular, let alone hard-bodied (i.e. shelled) organisms, like those found in the Cambrian strata. Bioherms, stromatolites, and other algal features are abundant, which Dr. Snelling notes is “hardly surprising” given that these sediments were not buried catastrophically. But are we to believe that not a single shell, tooth, scale, or anything else was preserved in sediments that were accumulating until the day of the Flood? If you’ve been to the beach, you’ll notice right away that shell fragments are littered throughout the coastal sands (and deeper water substrate if you care for a scuba-diving adventure). However, Dr. Snelling would have us believe that none of them were preserved in the young (<2,000 years) pre-Flood sediments, and that a majority were not even preserved in the Paleozoic sediments (Cambrian through Permian).
In conclusion, I believe that the Precambrian sedimentary rocks of the Grand Canyon (and many other parts of the world, for that matter) provide an insurmountable challenge to Flood geology. Thus I will devote a later article to this matter in particular. For now, I will continue under Dr. Snelling’s assertion that these sediments simply belonged to the pre-Flood ocean.
I say fossil, you say...fossil?
When I say the word fossil, what do you envision? Perhaps you have collected them yourself? If so, you’re probably familiar with trilobites, mollusks, ammonites, or even crinoids. Or, if you grew up in proximity to Wyoming, you have most certainly heard of the Green River basin, in which millions of pristine fish fossils have been recovered (in addition to crocodiles, turtles, plants, insects, and more). But I would dare say that our conception of fossils has been twisted to some degree Jurassic Park — yes, the movie that taught us all how to avoid being eaten by Tyrannosaurus and Velociraptor. Don’t get me wrong, I have nothing against museum displays of perfect body fossils, or movies about the recovery of ancient dinosaurs. However, all of the examples above are somewhat misleading about the nature of paleontology, in that they only show us fully articulated, well preserved fossils. Thus you would scarcely realize that a vast majority of recovered fossils (99%) are incomplete fragments of bones and shells, or individual teeth.
The reason I mention this is that it tends to contradict the Flood geology model. Granted, we may expect a global catastrophe to dismember many an unfortunate dinosaur, knock a few teeth loose, and break a number of shells. But if these critters were buried rapidly, then it seems unlikely that organic constituents (like muscle, tendons, and ligaments) could be sufficiently dissolved within days or months to leave almost nothing but disarticulated bones in the sediments. Even if this were the case, it would rule out survivability and hydrodynamic sorting as viable mechanisms for sorting a majority of fossils found. Again, I will grant the benefit of the doubt, but let us proceed with these principles in mind.
Fossils of the Grand Canyon and Grand Staircase
In his article, Dr. Snelling offers a rather simplified overview of fossils that have been found in the Grand Canyon strata. Now we shall ask the question, what do these fossils actually tell us?
 |  |
| Stromatolites from the late Cambrian of Nevada |
Nobody would be surprised that shallow marine invertebrates (trilobites, crinoids, brachiopods, etc.) are the first to show up in Cambrian rocks. This is consistent with evolutionary theory as well as the notion that a transgression of ocean water buried the first fossiliferous sediments over the Grand Canyon area. However, the complexity of biostratigraphy (matching layers based on their fossils) is hardly represented here. It’s not as though hundreds of species of trilobites, for example, are scattered throughout Paleozoic rocks in no particular order. On the contrary, particular assemblages of individual trilobite species (biomeres; Palmer, 1965) always show up in the same order. Imagine that you had documented 26 trilobite species throughout the world and assigned a letter to each one of them, depending on the order in which they were found. Now, in any given area, you might find a portion of the alphabet. It may not always be a complete alphabet (A, B, E, F, G, M, O, Q, Z), but it would always be found in the proper order. This is obviously not the result of any random process, but what reason do we have to expect this detail of order, assuming a Flood model? The order of trilobite species does not correlate to shape, size, shell chemistry, rock type, or any other characteristic that could be invoked in the Flood model. Thus we are left with trying to interpret the pattern in terms of original ecological zonation (i.e. maybe certain species lived further from the shore than others?). However, this hypothesis breaks down on three major points: 1) in that it requires nearly identical geography and hydrological forces on every continent during a catastrophic flooding of the world; 2) in that it must also account for species of brachiopods, mollusca, foraminifera, conodont, fish, and more, that are associated with the same assemblages of trilobite species; 3) it must also account for identical stratigraphic variations in chemical proxies (carbon, oxygen, and strontium isotope ratios, for example). Thus a careful transplant of detailed ecological niches must occur in the same order, everywhere on Earth, without significant mixing of sediments or water bodies.
Yet Dr. Snelling invokes the idea that repeated transgressions of marine water/sediments could account for the fact that terrestrial (land-based) sedimentary rocks are sandwiched between ‘marine’ successions. In other words, the Redwall Limestone and Kaibab Limestone contain only marine fossils, while the Coconino Sandstone contains none; the Dakota Sandstone and Straight Cliffs Formation are dominantly terrestrial, while the Tropic Shale is dominantly marine. What reason do we have to believe, however, that not a single shallow-marine trilobite survived to the deposition of the Tropic Shale, while deeper water ammonites did?
Footprints and vertebrate body fossils
Another major point of Dr. Snelling’s article is that vertebrate footprints are found in Paleozoic rocks, while fossils of their bodies are not found until higher up in the section. He interprets this as indicating that fleeing animals left tracks in the sediments, only to be overwhelmed and buried later on in the flood. While this seems plausible at first for the Grand Canyon sequence, it is simply not true when applied to other regions. Furthermore, fragments of reptile fossils have been found within the Supai Group (Harris et al., 1997), and more complete fossils are known from this time period (the Carboniferous) in other regions of the world (e.g. Müller and Reisz, 2005).
Regardless of Dr. Snelling’s interpretation of fossilized vertebrate trackways and body fossils, this phenomenon is not inconsistent with the conventional interpretation of Grand Canyon sedimentary rocks. Many tracks are present within the Coconino Sandstone, for example, while bones and teeth are notably absent. If the Coconino Sandstone represents deposition within a desert environment (like the Saharan desert), then it is not surprising that bones would be absent or very rare. In other words, there is a twofold preservation bias associated with a desert environment: 1) bones are less likely to be preserved in sediments, due to slow accumulation in an open, oxidizing environment, where life is scarce to begin with and depends on scavenging; 2) vertebrate fossil collection from cliff-forming sandstones is especially difficult (as opposed to silt/clay, which can be sifted), so it is premature to conclude that no body fossils are present in the formation.
Dinosaurs and mammals
Several summers ago, I had the opportunity of collecting thousands of fossil specimens from Bryce Canyon National Park, which were taken from the Straight Cliffs and Kaiparowits formations. A majority of the fossils were teeth, vertebrae, and scales from freshwater fish (including rays), and fragments of turtle shell. Also present were freshwater shark teeth, lizard, various amphibians, crocodiles, mammals, and of course, dinosaurs. The particular assemblage depended heavily on the type of sediment containing the fossils, which helped us to interpret the environment in which the animals lived. Moreover, fossils were not abundant throughout the rocks, but were typically confined to clay-rich mudstones typical of a floodplain environment, rather than in lenses of sandstones that represented infilled channels of ancient rivers.
Three lessons can be taken from my own experience that are relevant to the discussion. First, no distinctly marine fossils were found in sediments containing terrestrial fossils, while marine units did contain abundant marine fossils, but no trace of terrestrial organisms. This stratigraphic distinction is far too convenient for the Flood model, in which rapid transgression and burial would have been responsible for the alternation. Secondly, all fossils collected were unique to that time period, and the succession could be correlated with others around the world (for example, in Uzbekhistan, where another of our field assistants had done research). In other words, there were no remnants of trilobites, brachiopods, crinoids, earlier dinosaurs and other reptiles, amphibians, fish, sharks, or anything else that can be found lower in the Grand Canyon/Grand Staircase sequence. Third and finally, all mammals recovered from these sediments were represented by individual bones (particularly teeth), and were very small (less than 1 mm in width and height), while dinosaurs were represented by teeth (still small, less than 5 mm) and bone fragments (some small, some very large). Again, this is consistent with both the conventional geological interpretation, as well as evolutionary theory. Thus we are left to wonder why no traces of other mammals (cats, horses, monkeys, bats, megafauna) can be found in these sediments, while no dinosaurs are found in sediments further up, even though both groups come in all shapes and sizes. Even if we could accept that the differentiation is entirely due to distinct ecological niches or some kind of mammalian advantage in escaping the oncoming water, the principle does not apply to juveniles (which are better represented in the fossil record) or the recently deceased (did no mammals, or flowering plants for that matter, live/die near the shore?).
So it would seem that the most parsimonious interpretation of the order of fossils in the Grand Canyon and Grand Staircase sedimentary sequences is that they indeed represent assemblages of animals living at the respective time period. While the interpretations of Flood geologists can explain a general order in very limited cases, it does not hold up to scrutiny upon more detailed examination.
On the ‘predicted evolutionary order’ of fossils
I feel that Dr. Snelling is not as generous as I when it comes to allowing the opposing side to interpret the data. One example of this comes with the following statement:
‘Evolution predicts that new groups of creatures would have arisen in a specific order. But if you compare the order that these creatures first appear in the actual fossil record, as opposed to their theoretical first appearance in the predictions, then over 95% of the fossil record’s “order” can best be described as random. On the other hand, if these organisms were buried by the Flood waters...the major groups should appear in the fossil record according to where they lived, and not when they lived.’
It appears that Dr. Snelling is suggesting that evolutionary theory predicts new organisms to appear everywhere in the fossil record at the same time, regardless of depositional environment. Think of it this way. Imagine that scientists introduced a new kind of shallow marine arthropod to the Carribean (genetically engineered?) at a single point in time (let’s say next year). If we come back in a thousand years, would we expect to find remnants of the organism off the African coast, in the Persian Gulf, Caspian Sea, and desert dunes from California? Very possibly, we would find that the new organism migrated along the Carribean coast (maybe up the Carolina coast), but my point is that according to evolutionary theory also, the first appearance of a certain type of fossil in any given sequence of sedimentary rocks is controlled by where the animal lived, and not simply when. Thus it is unlikely that we should find a long, evolutionary succession of land plants or tetrapods (e.g. reptiles and amphibians) in the Grand Canyon, since a majority of sediments recorded during those time periods are from marine environments. When sections from around the world are compiled, however, the evolutionary history of life comes together very well. Not only can we document the first appearances of a given organism in the world, but we can often see how it migrated over time to other parts of the world.
Concluding thoughts
When I first began to research this article, I suspected that Dr. Snelling’s summary would hold up fairly well in the case of the Grand Canyon sediments, and that one would have to look in other sections of the world to falsify it. I grant that he is attempting to account for a massive amount of data (the fossil record) with a rather new and undeveloped model (Flood geology) in a specific case (the Grand Canyon). However, it is clear that the Flood model can not account for the order of fossils in the geologic record, even when applied to a small case in point. Moreover, only a fraction of geologic history is recorded within the Grand Canyon and Grand Staircase sedimentary sequences, and when we consider sections from other regions of the world, the conventional model remains consistent and predictive. The fossil record has been a long standing challenge to Flood geology, and I believe that it remains so for good reason.
References Cited:
Álvaro, J.J., and Debrenne, F., 2010, The Great Atlasian Reef Complex: An early Cambrian subtropical fringing belt that bordered West Gondwana: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 294, p. 120–132.
Austin, S.A., 1994, Grand Canyon: Monument to Catastrophe: Institute for Creation Research, 284 p.
Harris, A.G., Tuttle, E., Tuttle, S.D., 1997, Geology of National Parks (5th ed.): Kendall/Hunt Publishing.
Müller, J., and Reisz, R.R., 2005, An early Captorhinid reptile (Amniota, Eureptilia) from the Upper Carboniferous of Hamilton, Kansas: Journal of Vertebrate Paleontology, v. 25, p. 561-568.
Palmer, A.R., 1965, Biomere — a new kind of biostratigraphic unit: Journal of Paleontology, v. 39, p. 149-153.
Scherreiks, R., Bosence, D., BouDagher-Fadel, M., Melendez, G., Baumgartner, P.O., 2010, Evolution of the Pelagonian carbonate platform complex and the adjacent oceanic realm in response to plate tectonic forcing (Late Triassic and Jurassic), Evvoia, Greece: International Journal of Earth Science, v. 99, p. 1317-1334.
Sheehan, P.M., and Harris, M.T., 2004, Microbialite resurgence after the Late Ordovician extinction: Nature, v. 430, p. 75–78.
No comments:
Post a Comment